
Journal of Raptor Research Journal of Raptor Research
Volume 33 Issue 4 Article 1
January 2024
Population Density of Northern Spotted Owls in Managed Young-Population Density of Northern Spotted Owls in Managed Young-
growth Forests in Coastal Northern California growth Forests in Coastal Northern California
Lowell V. Diller
Darrin M. Thome
Follow this and additional works at: https://digitalcommons.usf.edu/jrr
Recommended Citation Recommended Citation
Diller, Lowell V. and Thome, Darrin M. (2024) "Population Density of Northern Spotted Owls in Managed
Young-growth Forests in Coastal Northern California,"
Journal of Raptor Research
: Vol. 33 : Iss. 4 , Article
1.
Available at: https://digitalcommons.usf.edu/jrr/vol33/iss4/1
This Scienti9c Paper is brought to you for free and open access by the Searchable Ornithological Research Archive
at Digital Commons @ University of South Florida. It has been accepted for inclusion in Journal of Raptor Research
by an authorized editor of Digital Commons @ University of South Florida. For more information, please contact

THE JOURNAL OF RAPTOR RESEARCH
A QUARTERLY PUBLICATION OF THE RAPTOR RESEARCH FOUNDATION, INC.
VOL. 33 DECEMBER 1999 NO. 4
j Raptor Res. 33(4):275-286
¸ 1999 The Raptor Research Foundation, Inc.
POPULATION DENSITY OF NORTHERN SPOTTED OWLS IN
MANAGED YOUNG-GROWTH FORESTS IN COASTAL
NORTHERN CALIFORNIA
LO•T•LL V. DILLER
Simpson Timber Company, P.O. Box 68, Korbel, CA 95550 U.&A.
DARRIN M. THOME
U.S. Fish and Wildlife Service, 2321 West Royal Palm Road, Suite 103, Phoenix, AZ 85021 U.S.A.
ABSTRACT.--We estimated population densities of Northern Spotted Owls (Strix occidentalis caurina) in
managed young-growth forests in coastal northern California from 1991-97. The 1266 km 2 study area
was divided into three subregions (Klamath--666 km 2, Korbel--392 km • and Mad River--208 km 2) and
completely surveyed each of the seven years. A total of 446 individual owls was marked to generate both
empirical and Jolly-Seber (J-S) estimates of density. Mean empirical and J-S estimates of abundance were
similar but mean estimates of crude density (territorial owls/km 2) differed among the three subregions
(Klamath--0.092 _+ 0.006 [_SE], Korbel--0.351 _+ 0.011, Mad River--0.313 _+ 0.017 and overall mean-
0.209 + 0.009). Significant differences in forest age-class composition among the three subregions
provided a plausible explanation for the low Klamath density but did not account for the similar den-
sities observed in -Korbel and Mad River. Ecological densities (number of individuals/area of habitat)
were higher than crude densities but the interpretation of this was limited because only nesting habitat
was used to estimate suitable habitat. Compared to limited published reports, densities were relatively
high in two of the three subregions in our study but this was probably typical of Northern Spotted Owl
densities for portions of coastal northern California. Recognizing the limitations of using density to
indicate habitat quality, our study provided valuable baseline data for assessing long-term trends in
Northern Spotted Owl population dynamics within the study area.
KEY WOP, DS: Northern Spotted Owl; Strix occidentalis caurina; California; density; managed forests; mark-
recapture.
Densidad poblacional de Strix Occidentalis caurina en los bosques jtvenes y manejados de las costas del
norte de California
RESUMEN.--Estimamos la densidad poblacional de Strix occidentalis caurina en bosques jtvenes y mane-
jados de las costas del norte de California entre 1991-97. Los 1266 km a del area de estudio fueron
divididos en tres subregiones (Klamath--666 km a, Korbel--392 km a y Mad River--208 km a) y los mon-
itoreamos durante los siete aftos. Un total de 446 individuos de buhos fueron marcados con el fin de
generar estimativos de densidad empiricos y de Jolly-Seber (J-S). La media empirica y los estimativos
de J-S de abundancia fueron similares, pero la media de densidad cruda (buhos territoriales/km a)
difiri6 en las tres subregiones (Klamath--0.092 + 0.006 [_SE], Korbel--0.351 _ 0.011, Mad River--
0.313 - 0.017 y la media promedio--0.209 + 0.009). Las diferencias significativas en la edad y clase de
la composicitn de los bosques entre las tres subregiones pueden ser la explicacitn de la baja densidad
de Klamath pero no para las densidades similares observadas en Korbel y Mad River. Las densidades
ecoltgicas (nfmero de individuos/area de habitat) fueron mayores que las densidades crudas. La in-
terpretacitn de esta fue limitada debido a que se utiliza el habitat de anidacitn para estimar habitats
convenientes. A1 comparar la limitada publicacitn de reportes, se encontr6 que las densidades fueron
275

276 DILLER AND THOME VOL. 33, NO. 4
relativamente altas en dos de las tres subregiones de nuestro estudio. Quizas esto sea tipico de las
densidades de Strix occidentalis caurina en porciones costeras del norte de California. AI reconocer las
limitaciones de usar densidades para indicar la calidad de habitat, nuestro estudio provee valiosos datos
para evaluar tendencias en el largo plazo sobre la din/unica poblacional de Strix occidentalis caurina
dentro del/trea de estudio.
[Traducci6n de Cfisar M/trquez]
The Northern Spotted Owl (Strix occidentalis
caurina) is associated with mature and old-growth
forests throughout much of its range. This rela-
tionship has been studied primarily through radio-
telemetry data that infers habitat selection through
disproportionate use of mature- and old-growth
forests relative to their occurrence within a land-
scape (Forsman et al. 1984, Carey et al. 1990, Solis
and Guti(•rrez 1990, Carey et al. 1992). In addition,
studies of Northern Spotted Owl occurrence and
abundance have shown a greater number of owl
sites in mature- and old-growth forests relative to
adjacent young forests (Forsman et al. 1977, Fors-
man et al. 1987, Forsman 1988, Bart and Forsman
1992, Blakesley et al. 1992). Given the economic
value of mature- and old-growth forests, the asso-
ciation of Northern Spotted Owls with these forests
places it at the center of a major controversy in the
Pacific Northwest. The 1990 listing of the Northern
Spotted Owl under the federal Endangered Spe-
cies Act (USDI 1992) instituted management pol-
icies limiting timber harvest of Northern Spotted
Owl habitat on public and private lands (Thomas
et al. 1990, Guti(•rrez et al. 1996, Marcot and
Thomas 1997).
The population density of a species is important
to resource managers for several reasons. In har-
vested game species, it is important to increase
population density to generate a greater harvest-
able surplus, and it may also be important to un-
derstand the population density relative to carry-
ing capacity (Krebs 1985, Caughley and Sinclair
1994). In species of conservation concern, popu-
lation density has been used as one of the indica-
tors of habitat quality (Forsman 1988, Thomas et
al. 1990, Bart and Forsman 1992), and one of the
criteria for establishing federally designated critical
habitat areas (USDI 1992). In many populations,
density has been used as a surrogate for knowing
vital rates of populations that allow estimation of
the population stability or viability.
Most attempts to compare abundance of North-
ern Spotted Owls in different habitats have relied
on estimates of relative abundance (Forsman et al.
1977, Marcot and Gardetto 1980), because esti-
mating population density has been difficult for a
species that exists in low numbers and occupies
large home ranges. As a result, reliable estimates
were not possible unless large areas were surveyed
(Franklin et al. 1990).
We surveyed 1266 km 2 of managed young-
growth forests for seven years as part of a monitor-
ing plan for the Northern Spotted Owl under
Simpson Timber Company's (STC) Habitat Con-
servation Plan (Simpson Timber Company 1992).
The primary objective of this study was to estimate
population density of owls in three subregions with
different forest age-class compositions to provide
baseline data for assessing long-term trends in
Northern Spotted Owl populations within a man-
aged young-growth landscape. We compared crude
(number of individuals/total area, Odum 1971)
and ecological densities (number of individuals/
area of habitat; Odum 1971), and assessed changes
in owl density during the study period (1991-97).
In addition, we compared estimates of abundance
based on empirical (direct counts of individuals for
which differences in detectability and sampling var-
iation associated with the estimate are not known)
and mark-recapture methods. Comparability of
these two approaches, empirical versus mark-recap-
ture, is important since most of the reported esti-
mates of Spotted Owl population density are based
on abundance estimates derived from empirical
data.
STUDY AREA
The study area was primarily within 1558 km e of land
owned by STC located in Del Norte, Humboldt and Trin-
ity counties, northwestern California. Most of this prop-
erty lies within 32 km of the coast, but can extend up to
85 km inland. The study area was located within the
Northern California Coast Range physiographic province
where fog is common (Mayer 1988). Near the coast,
mean summer and winter temperatures are about 18øC
and 5øC, respectively, whereas extremes of 38øC in sum-
mer and -IøC in winter are not uncommon beyond the
longitudinal belt of coastal influence approximately 48
km from the coast. Precipitation ranges from 102-254 cm
annually, with 90% of this falling from October-April (El-
ford 1974).
Predominate forest stands in the study area were coast-
al redwood (Sequ0ia sempervirens), Douglas-fir (Psendotsuga

DECEMBER 1999 NORTHErn SPOTTED OWL DENSITY 277
menziesii), and oak woodlands (Zinke 1988). Species char-
acterizing the oak woodlands included tanoak (Lithocar-
pus densiflorus), California black oak ( Quercus kelloggii)
and Oregon white oak (Q. garryana). Many of the red-
wood and Douglas-fir stands also contained a large com-
ponent of the following hardwoods: tanoak, bigleaf ma-
ple (Acer macrophyllum), madtone (Arbutus menziesii),
California bay ( Umbellular/a californica), and red alder (A1-
nus rubra).
Since the late 1960s, the primary silvicultural practice
has been even-aged management involving relatively
small clearcuts (12-24 ha in size) followed by prompt
replanting. About 97% of the study area consisted of
young forests ranging from 0-80 yr old. Residual trees
(left from past logging operations) were a component of
some forest stands and commonly the largest, oldest trees
present.
METHODS
Within STC lands, Northern Spotted Owl survey
boundaries were established apr/or/based on ownership
patterns, topographic features, vehicular access and oth-
er logistic considerations. The resulting study area was
further subdivided due to geographic and vegetative pat-
terns. In a nearby study area, Franklin et al. (1990) de-
termined that areas exceeding 90-130 km • were suffi-
cient to accurately estimate Northern Spotted Owl
density. Three subregions in our study area met this cri-
terion and hereafter are referred to as Klamath (666
kmg), Korbel (392 km 9) and Mad River (208 kmg; Fig.
1). Other isolated tracts of STC property were too small
to be included as separate subregions. Following Thome
et al. (1999), we created six categories of stand age to
classify habitat: 0-5, 6-20, 21-40, 41-60, 61-80, and •80
yr (Table 1). The 61-80 and •80 yr age classes were com-
bined for this analysis, because there was very little area
of one or both of these age classes in the three subre-
gions.
We surveyed the entire STC study area for Northern
Spotted Owls at least twice each season using a complete
and systematic search protocol from I March-30 August,
1991-97. Prior to initiation of surveys, we inspected the
entire study area using 1:24000 aerial photographs. We
plotted call points at strategic locations that maximized
observer ability to solicit and detect responses from owls.
Call points were usually positioned at relatively high el-
evations with unobstructed forest openings to ensure a
clear and far-ranging broadcast of the call. Solicitations
consisted of playing recorded Northern Spotted Owl calls
or vocalizing imitations of calls for a minimum duration
of 10 min. We used a jet boat to access and survey STC
property bordering the Klamath River. All surveys using
this protocol were conducted nocturnally, beginning no
earlier than dusk. If an owl responded to a nocturnal call,
•ts location was plotted, and a daytime follow up effort
was initiated, where an observer attempted to locate the
roosting owl by pursuing responses made to imitated or
recorded calls (Forsman 1983). We captured owls using
noose or snare poles (Forsman 1983) and banded them
with a USGS band on one leg and a plastic, color-coded
band on the other (serving as a unique identifying mark;
Forsman et al. 1996). Sex and age were determined fol-
lowing Forsman (1981, 1983) and Moen et al. (1991).
We calculated forest stand ages using STC's timber •n-
ventory database in Intergraph's CAD system, integrated
with the Modular Graphics Environment 5.0 (Intergraph
Corporation 1994) geographic information system (GIS).
Forest stands were distinguished based on date of harvest
and polygons were drawn around unique forest stands.
Only GIS data from 1997 were available for analysis
Landscape data from 1997 were considered adequate be-
cause the mean annual percent change in the landscape
(from timber harvest) during this study was 0.7 ñ 0.08
[ñSE], 1.0 _ 0.18 and 0.5 ñ 0.16% for the Klamath,
Korbel and Mad River study areas, respectively.
Not all of the land surveyed was owned by STC, be-
cause other private lands (in-holdings) were common
within our study area, and survey boundaries were set by
topographic features and access points rather than own-
ership boundaries. Since GIS coverage was limited to
STC lands, we were able to assess age-class conditions for
90% (599 km 9) of Klamath, 75% (294 km •) of Korbel
and 70% (145 km 9) of Mad River. Despite this, we believe
the GIS coverage was representative of the entire study
area, since most of the landscape was subjected to the
same historic timber harvesting practices that created en-
tire watersheds with similar aged stands. In addition, the
in-holdings and adjacent lands associated with the Korbel
and Mad River subregions (areas with the least GIS cov-
erage) were virtually all private lands zoned for timber
production. We compared the amount of forest in the
five age classes among the three subregions (Table 2)
using Chi-square analysis (Hintze 1997).
We used the Jolly-Seber (J-S) capture-recapture model
(Jolly 1965, Seber 1965, 1982) that allowed for death and
immigration in open populations. We used program JOL-
LY (Pollock et al. 1990) to calculate J-S estimates of an-
nual abundance (Nt). Because population and density es-
timates on STC lands had never been documented, we
were primarily interested in these parameters from the
modeling. We subjectively chose the reduced parameter
J-S model (model D in program JOLLY) to analyze the
data, because reduced parameter models compute abun-
dance estimates with greater precision than models sat-
urated with parameters (Jolly 1982). Ninety-five percent
confidence intervals were calculated as 1.96 (SE [Nt]).
Goodness-of-fit tests (Pollock et al. 1985) in program
JOLLYwere used to determine if the models fit the data.
When goodness-of-fit tests suggested lack of fit, we used
a variance inflation factor, •, based on quasi-likelihood
theory (Burnham et al. 1987:243-246, McCullagh and
Nelder 1989) to adjust variances in models with overdis-
persed data (Lebreton et al. 1992, Anderson et al. 1994).
The variance inflation factor is calculated as Xg/V where
X 9 was the goodness-of-fit statistic with v degrees of free-
dom. Expected values for • are not, on average, different
from 1.0 with models that fit the data, and do not exceed
•4 in models that attain structural adequacy, but may
need variance inflation measures (values of 6-10 indicate
complete model inadequacy requiring an entirely new
model). If • indicated that variance inflation measures
were necessary, the standard error of each population
parameter was calculated as X/•SE} (Anderson et al.
1994).
Empirical estimates of annual abundance (Nt) fol-
lowed criteria established in Franklin et al. (1990), which

278 DILLER AND THOME VOL. 33, NO. 4
STUDY AREA
CRESC•ENT Ci'TY
,-•- CALIFORNIA
",,, k
SPOTTED OWL TERRITORIES
Figure 1. Map of the Simpson Timber Company study area, northwest California. Dots represent Northern Spotted
Owl locations within and adjacent to Klamath, Korbel and Mad River subregion boundaries.

DECEMBER 1999 NORTHERN SPOTTED OWL DENSITY 279
Table 1. Description of six forest age categories used in analysis of Northern Spotted Owl ecological density for the
Simpson Timber Company study area in northern California, 1991-97.
TREES/ha BASAL AREA a VOLUME b
AGE
CATEGORY • SD i SD • SD
0-5 0.9 5.9 0.2 1.0 0.1 0.7
6-20 42.2 160.8 2.3 8.4 0.8 4.3
21-40 558.6 292.6 29.7 15.8 6.7 7.4
41-60 708.2 320.9 46.9 18.5 14.6 11.2
61-80 591.4 384.9 59.1 18.3 29.8 19.8
>80 811.6 598.9 58.4 30.7 28.7 27.8
m2/ha.
Million board m/ha.
assumed an annual census of territorial owls in which all
individuals known to be alive in the study area were
counted. The total annual count was based on surveys
over the 7-yr period and included the: number of iden-
tified (banded) individuals; number of unidentified in-
dividuals mated to identified owls; and number of un-
identified individuals assumed different from identified
individuals in nearby territories.
Population density was estimated as crude density (N t
/total area; Odum 1971) and ecological density (N t/area
of habitat; Odum 1971). We used J-S estimates of adult
and subadult Northern Spotted Owls within the three
subregions for N t. Following the rationale of Franklin et
al. (1990), we used the estimated total quantity of North-
ern Spotted Owl habitat as the divisor to calculate eco-
logical densities. In their study, the proportion of telem-
etry locations of owls in different habitats was used as one
method to estimate total owl habitat. Old-growth, which
had the highest proportion of telemetry locations, was
assigned a weight of 1.0 with other habitats weighted
based on the proportion of telemetry locations in those
habitats relative to those in old-growth (Franklin et al.
1990). Since we had no telemetry data to assess foraging
habitat in our study area, we calculated the total owl hab-
itat in each subregion based on the relative amount of
nesting habitat.
To calculate ecological densities, we assigned a weight
of 1.0 for the >60 yr age class, because it had the highest
proportion of nest sites relative to the total forested area
in the age class (0.27 nests/km2). Other age classes were
then weighted (normalized) by dividing the proportion
of nest sites in those age classes by the proportion of
nests in the >60 yr old age class (Table 3). For example,
there were 0.18 nests/km 9 in the 41-60 yr age class,
which was 68.5% of the density found in the >60 yr old
age class. Crude densities were calculated as N t (J-S) di-
vided by the size of the associated subregion. Ninety-five
percent confidence intervals for the density estimates
were calculated by dividing the population confidence
intervals by the subregion size (Seber 1982).
Abundance estimates cannot be computed for the im-
tial year of study using program JOLLY. Therefore, we
used preliminary capture data from 1990 as the first year
of analyses, even though a complete census protocol was
not established until 1991 (Franklin et al. 1990). In 1990,
we banded and subsequently entered in the analysis, 14,
76 and 17 owls from Klamath, Korbel, and Mad River,
respectively. We used program CONTRAST (Hines and
Sauer 1989) to examine differences in abundance esn-
mates among years for the three subregions. Program
CONTRAST uses a general Chi-square statistic to test dif-
ferences among abundance estimates using contrasts
(Sauer and Williams 1989). We first tested for overall ho-
mogeneity in abundance estimates for each subregion. If
a test yielded significant results, we then tested a-postenort
to determine which years were causing heterogeneity. Al-
pha levels for a-posteriori tests were adjusted to maintain
the overall experiment-wise error rate (Neter and Was-
serman 1974). We used the Bonferroni approach of us-
ing ot/m as the significance level for unplanned compar-
Table 2. Percent of forest habitat in five age classes and percent of nonforest on three subregions of the Simpson
Timber Company (STC) study area in northern California.
FOREST AGE CLASS IN YEARS
SUBREGION 0--5 6--20 21--40 41--60 >61 NONFOREST NONSTC a
Klamath 4.3 27.9 49.4 5.2 10.6 2.6 10.0
Korbel 6.0 24.8 31.2 24.3 10.1 3.5 25.0
Mad River 3.6 3.8 16.0 23.8 34.2 18.6 30.0
Percent of total study area within each subregion that was not within STC ownership or for which there was no forest age class
•nformation.

280 DILLER AND THOME VOL. 33, NO. 4
Table 3. Habitat weight and amount of weighted habitat
in each age class for three subregions of the Simpson
Timber Company study area in northern California. Hab-
itat weights were calculated from 86 nest sites of North-
ern Spotted Owls, 1991-97.
FOREST AGE CLASS IN YEARS
0-5 6-20 21-40 41-60 >61 TOTAL a
Habitat weight b 0 0.02 0.20 0.68 1.00
Amount of weighted habitat c by subregion (km 2)
Klamath 0 2.8 66.4 23.8 70.8
Korbel 0 1.4 24.7 65.3 39.6
Mad River 0 0.1 6.7 33.9 71.1
163.7
131.1
111.9
Excludes nonforested areas.
See methods for description of approach used.
Amount of forest multiplied by habitat weight.
isons, where m was the number of unplanned tests. All
tests were performed with a significance level of 0.05.
RESULTS
A Chi-square analysis indicated that there was a
significant difference in forest age-class composi-
tion among subregions (X 2 -- 201.30, df = 8, P <
0.001; Table 2). Klamath had the highest propor-
tion of stands in younger age classes (83.7% <40
yr old) followed by Korbel (64.3% <40 yr old) and
Mad River (28.7% <40 yr old).
A total of 103, 228 and 115 adult and subadult
Northern Spotted Owls were banded at 55, 80 and
47 territories in the Klamath, Korbel and Mad Riv-
er study areas, respectively, from 1990-97 (Fig. 1).
Estimates of capture and survival probabilities were
generally high and were similar among all three
study areas (Table 4). The J-S model fit the data
well for Klamath (X 2 = 19.51, df = 18, P = 0.361),
but not for Korbel (X 2 = 89.37, df = 24, P< 0.001)
and Mad River (X 2 -- 54.91, df = 18, P < 0.001).
We used variance inflation factors for Korbel (• =
3.72) and Mad River (• = 3.05) to adjust the sam-
pling variance of the abundance estimates.
Abundance estimates appeared to increase over
the first two years of the study (Fig. 2). The overall
test of homogeneity for abundance estimates over
the seven years yielded significant differences for
Klamath (X 2 = 22.80, df = 6, P < 0.001), Korbel
(X 2 -- 27.49, df = 6, P < 0.001) and Mad River (X 2
= 14.14, df = 6, P = 0.028). The 1991 abundance
estimates for Klamath (48.91 -+ 3.65 [+-SE]) and
Korbel (117.24 +- 6.62) were significantly lower
than their mean estimates for the other years,
Table 4. Jolly-Seber estimates of capture probabilities
(P), apparent survival probabilities (qb) and percent co-
efficient of variation (CV) for mean abundance estimates
of Northern Spotted Owls for three subregions of the
Simpson Timber Company study area in northern Cali-
fornia, 1991-97.
SUBREGION P SE (P) qb SE (qb) CV (%)
Klamath 0.78 0.03 0.87 0.02 6.7
Korbel 0.84 0.01 0.88 0.01 3.1
Mad River 0.82 0.02 0.85 0.02 5.6
1992-97 (Klamath: • = 63.09 i 1.23; X 2 = 13.56,
df = 1, P < 0.001; and Korbel: i = 140.81 i 2.69;
X 2 = 10.88, df = 1, P = 0.001). The Mad River
abundance estimate for 1994 (78.50 i 4.67) was
significantly different from the mean estimate for
the other years (• = 62.82 -+ 2.00; X 2 = 9.52, df =
1, P = 0.002).'Bonferroni adjustments of the alpha
level prevented identifying additional significant
differences.
Empirical and J-S estimates of abundance
showed similar general trends for all subregions,
but there were some differences in individual es-
timates among some years. The confidence inter-
vals for J-S estimates did nbt overlap empirical es-
timates of the abundance during 1992-94, 1993-
96 and 1995 for Klamath, Korbel and Mad River,
respectively (Fig. 2). The mean empirical and J-S
estimates of abundance (Table 5) differed for Kor-
bel (X 2 = 6.805, df = 1, P = 0.009), but were not
significantly different for Klamath (X 2 -- 0.623, df
= 1, P = 0.430) or Mad River (X 2 = 0.792, df = 1,
P-- 0.373).
Mean J-S crude densities were highest for Korbel
followed by Mad River and Klamath (Table 5) with
an overall mean of 0.209 owls/km 2 (95% C.I. =
0.190-0.228). Ecological densities followed the
same trend as crude densities (Fig. 2) but calculat-
ed values were higher (Table 5). Comparisons of
mean crude and ecological densities indicated that
the three subregions were significantly different
for both variables (X 2 = 2038.098, df = 2, P <
0.001 and X •= 1249.670, df = 2, P < 0.001 for the
crude and ecological comparisons, respectively).
Post hoc comparisons showed crude and ecological
density estimates for all subregions to be different
from each other (Table 5, ecological densities: Kor-
bel vs. Klamath; X 2 = 4871.43, df = 1, P < 0.001;
Korbel vs. Mad River; X 2 = 38.35, df -- 1, P< 0.001;
Klamath vs. Mad River; X• = 1679.44, df = 1, P <
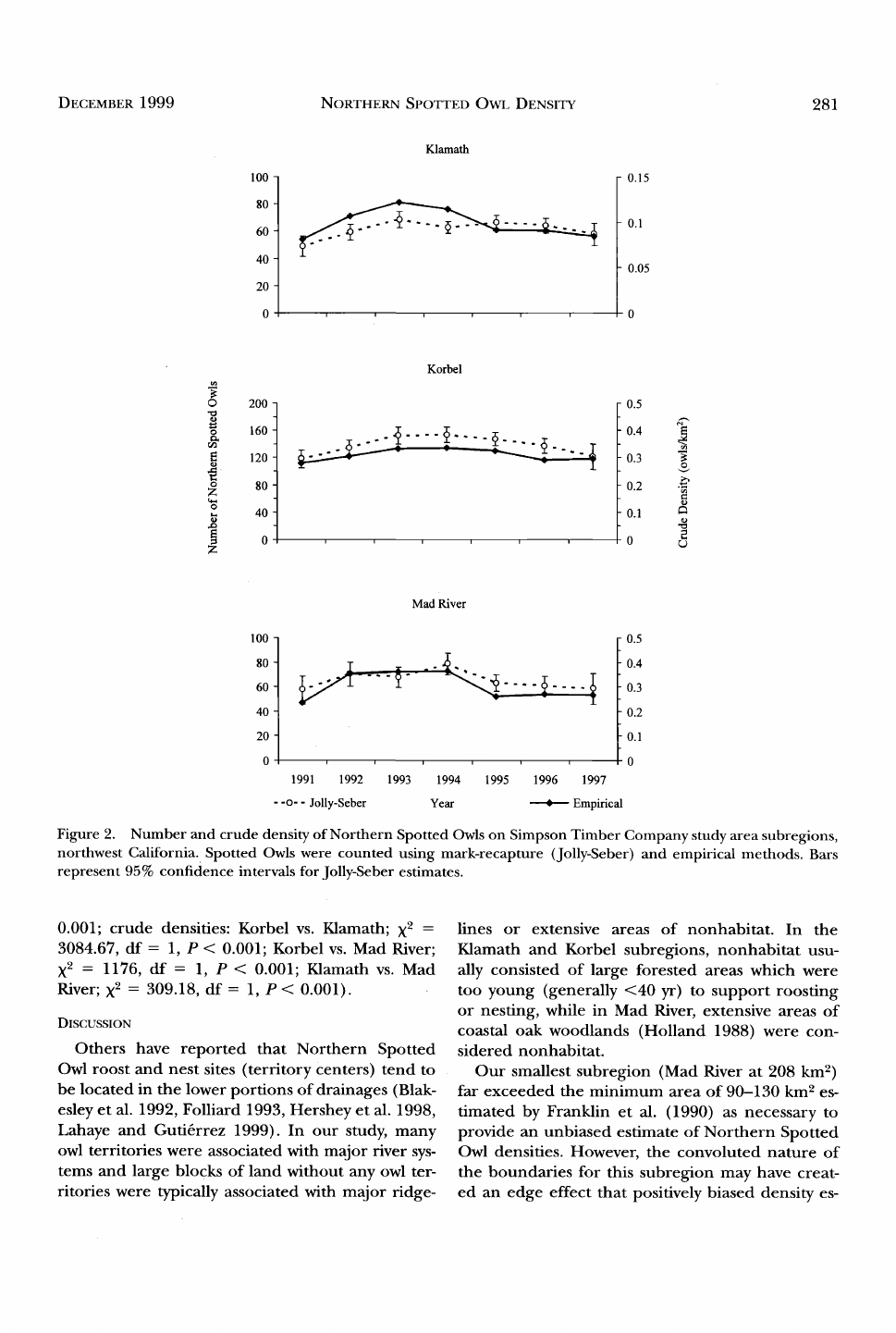
DECEMBER 1999 NORTHEed,• SPOTTED OWL DENSITY 281
lOO
80
60
40
20
o
Klamath
0.15
0.1
0.05
200
160
120
8O
40
0
Korbel
0.5
0.4
0.3
0.2
0.1
0
Mad River
lOO
8o
6o
40
20
o
1991 1992 1993 1994 1995 1996 1997
--o-- Jolly-Seber Year * Empirical
0.5
0.4
0.3
0.2
0.1
0
Figure 2. Number and crude density of Northern Spotted Owls on Simpson Timber Company study area subregions,
northwest California. Spotted Owls were counted using mark-recapture (Jolly-Seber) and empirical methods. Bars
represent 95% confidence intervals for Jolly-Seber estimates.
0.001; crude densities: Korbel vs. Klamath; X 2 =
3084.67, df = 1, P < 0.001; Korbel vs. Mad River;
X 2 = 1176, df = 1, P < 0.001; Klamath vs. Mad
River; X 2 = 309.18, df = 1, P < 0.001).
DISCUSSION
Others have reported that Northern Spotted
Owl roost and nest sites (territory centers) tend to
be located in the lower portions of drainages (Blak-
esley et al. 1992, Folliard 1993, Hershey et al. 1998,
Lahaye and Gutifirrez 1999). In our study, many
owl territories were associated with major river sys-
tems and large blocks of land without any owl ter-
ritories were typically associated with major ridge-
lines or extensive areas of nonhabitat. In the
Klamath and Korbel subregions, nonhabitat usu-
ally consisted of large forested areas which were
too young (generally <40 yr) to support roosting
or nesting, while in Mad River, extensive areas of
coastal oak woodlands (Holland 1988) were con-
sidered nonhabitat.
Our smallest subregion (Mad River at 208 km 2)
far exceeded the minimum area of 90-130 km • es-
timated by Franklin et al. (1990) as necessary to
provide an unbiased estimate of Northern Spotted
Owl densities. However, the convoluted nature of
the boundaries for this subregion may have creat-
ed an edge effect that positively biased density es-

282 DtLLER AND THOME VOL. 33, NO. 4
Table 5. Mean empirical and Jolly-Seber (J-S) estimates of Northern Spotted Owl abundance along with estimated
crude and ecological densities for three study area subregions of the Simpson Timber Company study area in north-
ern California, 1991-97.
ABUNDANCE ESTIMATES DENSITY ESTIMATES (owus/k•n 2)
EMPIRICAL J-S CRUDE a ECOLOGICAL b
SUBREGION • ----- SE • _+ SE • _+ SE / +-- SE
Klamath 65.6A c 3.95 61.1A 4.12 0.092A d 0 0.373A 0.015
Korbel 123.3A 3.36 137.4B 4.26 0.351B 0.014 1.049B 0.041
Mad River 60.1A 4.17 65.1A 3.63 0.313C 0.014 0.581C 0.026
aJ-S estimates used as the dividend to calculate number of owls/total area (Odum 1971).
bJ-S estimates used as the dividend to calculate number of owls/area of habitat (Odum 1971).
Means within rows and within abundance estimates followed by the same letter do not differ (P > 0.05).
Means within columns and within density estimates fbllowed by the same letter do not differ (P > 0.05).
timates. The other subregions (Klamath at 666 km '•
and Korbel at 392 km 2) were large enough that
edge effects should not have been a factor.
This study was patterned after the Northern
Spotted Owl density study by Franklin et al. (1990)
in the Willow Creek study area (WCSA) immedi-
ately to the east of the Mad River subregion. They
concluded that, because of the high capture and
survival probabilities and the corroborative evi-
dence provided by the empirical estimates, the J-S
model provided both an accurate and precise es-
timate of Northern Spotted Owl density. We also
found close agreement between the J-S and empir-
ical estimates, indicating that our estimates were
also accurate. The empirical estimate did signifi-
cantly underestimate density relative to J-S for the
Korbel subregion, but the magnitude of the differ-
ence was only 10.3%. The mean capture probabil-
ity in our study area (/= 0.81 _ 0.02) was lower
than that observed in the WCSA (J-S model D, •
= 0.91 -+ 0.30; Franklin et al. 1990), but compari-
sons using program CONTRAST showed no statis-
tical differences between the two study areas (X '• =
0.103, df = 1, P = 0.748). Comparison of mean
survival probabilities between the WCSA (• -- 0.89
+ 0.02; Franklin et al. 1990) and our study area (i
-- 0.87 + 0.01) also showed no difference (X •=
1.197, df = 1, P-- 0.274).
The apparent increasing abundance trend over
the first few years in all subregions was most likely
related to increased cumulative sampling effort
and not a real increase in abundance. Despite our
attempt to survey the entire study area each year,
some resident owls apparently were not located un-
til the second or even third year of the study. This
conclusion was based on the observation that many
of these newly discovered owls were adult breeding
pairs. If the newly discovered sites had resulted
from new birds that colonized sites subsequent to
the start of the study, they would most likely have
been nonbreeding subadult owls. Other owls were
missed in areas not surveyed in the early years of
the study because they were assumed to be non-
habitat but were subsequently found to contain
owls.
Similar to findings reported by Franklin et al.
(1990), we noted a close agreement between J-S
and empirical estimates. Mean absolute differences
between J-S and empirical abundance estimates
were only 10.3, 7.6 and 7.4% for Korbel, Mad River
and Klamath, respectively. The results of both stud-
ies could be interpreted to indicate that reliable
estimates of abundance (density) can be obtained
through empirical estimates without the effort and
cost associated with marking and recapturing birds
to obtain J-S estimates. However, we believe that if
a large portion of the population is unmarked, em-
pirical estimates would likely vary substantially due
to the high potential for "double counting" indi-
viduals in some situations and discounting new
birds in other circumstances. In addition, mean-
ingful comparisons among years or study areas
would be problematic because empirical estimates
do not account for differences in detectability or
sampling variation.
Our crude density estimates for the three sub-
regions (Klamath--0.092 owls/km2; Korbel--0.351
owls/km'•; and Mad River--0.313 owls/km '•) span
the reported ranges of population density for both
the Northern Spotted Owl and the California Spot-
ted Owl (S. o. occidentalis). Marcot and Gardetto
(1980) reported the equivalent of approximately

DECEMBER 1999 NORTHERN SPOTTED OWL DENSITY 283
0.325 owls/km '• in the Six Rivers National Forest
which is similar to our estimates for Korbel and
Mad River. However, as noted by Franklin et al.
(1990), their estimate was based on empirical
counts from night surveys without marking birds,
and their largest study area was only 58.2 km 2. Both
of these factors would likely positively bias their es-
timates making comparisons to this study problem-
atic. The lower population density in Klamath is
similar to many of the reported densities of Cali-
fornia Spotted Owls in the Sierra and San Berna-
dino Mountains (Roberts 1993, Moen and Guti6r-
rez 1993, Lahaye and Gutierrez 1994). Franklin et
al. (1990) provided the most rigorous estimate re-
ported for the population density of Northern
Spotted Owls. They estimated a density of 0.235
owls/km 2 for the 292 km '• WCSA, which was inter-
mediate in study area size between the Korbel and
Mad River subregions of our study. Their estimate
was similar to our combined estimate (0.209 owls/
km2), but less than either Korbel or Mad River,
which were located in closest proximity to the
WCSA. Tanner and Guti6rrez (1995) estimated
0.219 owls/km = for a 137.7 km '• study area in Red-
wood National Park, which was the only previous
estimate of density for Northern Spotted Owls in
the coastal redwood region. This was an empirical
estimate based on two years of surveys, but most
owls were marked, thus the estimate was likely ac-
curate.
Without other density studies in the coastal red-
wood region of Northern California, it is difficult
to know the extent to which this study is represen-
tative of the region. However, we believe the pat-
tern of density we observed was reflective of the
region in general. This was based on a qualitative
assessment we conducted using the 1996 California
Natural Diversity Database (G. Gould, California
Department of Fish and Game, unpubl. data) of
reported Northern Spotted Owl locations across
the entire range of the subspecies in California
and on unpublished data from an adjacent large
industrial land owner (S. Chinnici, Pacific Lumber
Company, pers. comm.).
There was a significant difference in the amount
of forested habitat in specific age classes among
the three subregions. We could only speculate on
how this might have influenced owl density since
the study was not designed to assess this. Although
some young stands (20-40 yr) in the STC study
area were associated with high Northern Spotted
Owl fecundity and low turnover rates, forests <40
yr old were not selected in proportion to their
availability by owls for nesting (Thorne et al. 1999).
Thus, high proportions of stands <40 yr old might
limit owl density. Klamath had significantly lower
densities of owls than the other subregions along
with the highest proportion of the landscape m
younger stands (83.7% <40 yr old). Klamath also
tended to have extensive areas of homogeneous
younger age classes, although we have not quanu-
fled this difference. In comparison, Korbel had
high densities of owls, with 64.3% of forest stands
<40 yr old. Based on extensive harvesting in the
last 10-15 yr with relatively small clearcuts (10-24
ha), Korbel tended to have a much more hetero-
geneous mixture of stand ages relative to Klamath.
In the same study area, Folliard (1993) noted that
landscapes supporting Northern Spotted Owls had
more edge and greater stand diversity than ran-
domly selected landscapes. Finally, like Korbel,
Mad River had high densities of owls, but only
28.7% of stands were <40 yr old. We had no data
to establish a direct cause and effect relationship
between habitat variables and the density of owls
in the different subregions and comparing density
to habitat variables was not the primary objective
of this study. However, as noted by Thome et al.
(1999), a combination of different age classes (old-
er stands for nesting and younger stands for for-
aging) may provide the best habitat for Northern
Spotted Owls in our region.
By definition, ecological densities are equal to or
greater than crude densities, and one can predict
that the magnitude of the difference will increase
as the proportion of habitat for a given species de-
creases on the landscape. Ecological densities were
4.05, 2.99 and 1.86 times higher than crude densi-
ties for Klamath, Korbel and Mad River, respectively,
which supported the predicted differences based on
the relative amounts of habitat in each region. In
comparison, Franklin et al. (1990) reported ecolog-
ical densities that were 2.81 and 2.31 times higher
than crude densities depending upon the approach
used for defining owl habitat.
It is difficult to make meaningful comparisons
of ecological densities among studies in different
areas unless the same criteria are used to calculate
ecological densities. Using mature/old-growth for-
ests to represent owl habitat, Franklin et al. (1990)
reported an ecological density of 0.660 owls/km '•
in the WCSA. Their estimate of ecological density
was greater than our estimate for the Klamath re-
gion (0.373 owls/km=), less than Korbel (1.049

284 D•LL}•R AND THOME VOL. 33, NO. 4
owls/km 2) but quite similar to Mad River (0.581
owls/km2). In addition to being closest in prox-
imity to the WCSA, Mad River also had the highest
proportion of mature stands (36.9% >80 yr in age,
although it lacked old growth habitat) compared
to 35.6% mature/old growth in the WCSA.
There is some question as to the extent compar-
isons of Northern Spotted Owl densities, either
w•thin or between study areas, can be used for de-
veloping management prescriptions. As noted by
Van Horne (1983), population density of a species
can be a misleading indicator of habitat quality.
Although some of the attributes of Northern Spot-
ted Owl populations do not meet the criteria for
habitat quality-density decoupling, a prediction
consistent with decoupling habitat quality and den-
sity is that high owl densities on selected managed
lands result from displacement of owls from adja-
cent harvested areas. However, we believe this was
unlikely because the densities in our study area ap-
peared to be relatively stable throughout a time
period when, due to its federally-listed status
(USDI 1992), significant habitat alteration of
Northern Spotted Owl habitat was not permitted
on adjacent private lands. In addition, there was a
90-95% reduction in annual timber harvest on ad-
jacent public land (Six Rivers National Forest) just
prior to and after the listing of the Northern Spot-
ted Owl (USDA 1995). Finally, we have observed
that the highest reproduction tends to be associ-
ated with areas of highest densities (L. Diller, un-
publ. data), but it was beyond the scope of this
study to quantify the relationship between repro-
duction and density.
Although it was unlikely that the densities of
owls in our study area were influenced by displace-
ment from adjacent areas, we could not assess hab-
itat quality in our study area based on density of
owls. First and foremost, we could not establish
causal relationships between the observed differ-
ences in density and corresponding differences in
habitat attributes without undertaking an experi-
mental approach over large areas. Correlative stud-
ies to elucidate patterns between habitat attributes
and density were not possible when only a few sub-
regions were available for comparison. In addition,
we could only estimate the density of the territorial
population of owls, and true density, which would
include nonterritorial floaters, was unknown. Giv-
en the difficulty of undertaking experiments with
a protected species over large areas, we believe that
more immediate insight can be gained concerning
habitat quality by relating demographic parameters
to habitat attributes in a manner described in
Thome et al. (1999). Ultimately, knowing popula-
tion density is of limited immediate benefit for de-
veloping conservation strategies for Northern
Spotted Owls without knowing the habitat attri-
butes that result in demographic parameters that
will sustain populations over time. However, estab-
lishing reliable estimates of population densities
for Northern Spotted Owls should provide valu-
able baseline data for assessing long-term trends in
their populations. Similar studies should be con-
ducted in selected areas throughout the range of
the Northern Spotted Owl to allow future assess-
ment of the long-term response of this species to
current management strategies now being imple-
mented.
ACKNOWLEDGMENTS
We thankJ.M. Beck, G.H. Brooks, D.E. Copeland, L.B
Folliard, K.H. Hamm, CJ. Hibbard, R.R. Klug, B.D. Mi-
chaels, D.B. Perry, J.L. Thompson, J.P. Verschuyl and BJ.
Yost for their exceptional fieldwork. G.N. Warinner and
C.J. Lane were very patient in helping with STC's GIS
data. K.H. Hamm provided valuable editorial review and
data analysis for the development of the manuscript. An
ß earlier draft was greatly improved by comments fromJ.B.
Buchanan, L.L. Irwin and an anonymous reviewer.
L•TERATURr C•TEr)
ANDERSON, D.R., K.P. BURNHAM AND G.C. WHITE. 1994.
AIC model selection in overdispersed capture-recap-
ture data. Ecology 75:1780-1793.
BART, J. AND E.D. FORSMAN. 1992. Dependence of North-
ern Spotted Owls Strix ocddentalis caurina on old-
growth forests in the western USA. Biol. Cons. 62:95-
100.
BLAKESLE¾, J.A., A.B. FRANKLIN AND RJ. GUTieRReZ. 1992.
Spotted Owl roost and nest-site selection in north-
western California. J. Wildl. Manage. 56:388-392.
BURNHAM, K.P., D.R. ANDERSON, G.C. WHITE, C. BROWNIE
AND K.H. POLLOCK. 1987. Design and analysis meth-
ods for fish survival experiments based on release-re-
capture. Am. Fish. Soc. Monogx 5.
CAR•¾, A.B., J.A. RE•D AND S.P. HORTON. 1990. Spotted
Owl home range and habitat use in southern Oregon
Coast Ranges. J. Wildl. Manage. 54:11-17.
--, S.P. HORTON AND B.L. B•SWELL. 1992. Northern
Spotted Owls: influence of prey base and landscape
character. Ecol. Monogx 62:223-250.
CAUGHLEY, C-. AND A. S•NCLAIR. 1994. Wildlife ecology and
management. Blackwell Scientific Publications, Cam-
bridge, MA U.S.A.
ELFOre), R.C. 1974. Climate of Humboldt and Del Norte
counties. Humboldt and Del Norte Counties Agric.
Exten. Serv., Univ. California, Davis, CA U.S.A.

DECEMBER 1999 NORTHERN SPOTTED OWL DENSITY 285
FOLLIARD, L.B. 1993. Nest-site characteristics of Northern
Spotted Owls in managed forests of northwest Cali-
fornia. M.S. thesis, Univ. Idaho, Moscow, ID U.S.A.
FORSMAN, E.D. 1981. Molt of the Spotted Owl. Auk 98:
735-742.
1983. Methods and materials for locating and
studying Spotted Owls. USDA For. Serv. Gen. Tech.
Rep. PNW-162, Portland, OR U.S.A.
ß 1988. A survey of Spotted Owls in young forests
in the northern coast range of Oregon. Murrelet 69:
65-68.
, E.C. MESLOW AND MJ. S•:RUB. 1977. Spotted Owl
abundance in young versus old-growth forests. Oreg.
Wildl. Soc. Bull. 5:43-47.
--, --AND --. 1984. Distribution and biol-
ogy of the Spotted Owl in Oregon. Wildl. Monog½. 87:
1-64.
, C.R. BRUCE, M.A. WALKER AND E.C. MESLOW.
1987. A current assessment of the Spotted Owl pop-
ulation in Oregon. Murrelet 68:51-54.
, A.B. FRANKLIN, F.M. OLIVER AND J.P. WARD. 1996.
A color band for Spotted Owls. J. Field Ornithol. 67:
507-510.
FRANKLIN, A.B., J.P. WArn), RJ. GUTII•RREZ AND G.I.
GOULD, JR. 1990. Density of Northern Spotted Owls
in northwest California. J. Wildl. Manage. 54:1-10.
GUTIERREZ, RJ., E.D. FORSMAN, A.B. FRANKLIN AND E.C.
MESLOW. 1996, History of demographic studies in the
management of the Northern Spotted Owl. Stud. Ari-
an Biol. 17:6-11.
HERSHEY, K.T, E.C. MESLOW AND F.L. P,•USEY. 1998. Char-
acteristics of ibrests at Spotted Owl nest sites in the
Pacific Northwest. J. Wildl. Manage. 62:1398-1410.
HINES, J.W. AND J.R. S^UER. 1989. Program CONTRAST--
A general program for the analysis of several survival
or recovery rate estimates. USDI Fish and Wildl. Serv.,
Tech. Rep. 24. Washington, DC U.S.A.
HIN•:ZE, J.L. 1997. NCSS 97: Statistical system for win-
dows. NCSS, Kaysville, UT U.S.A.
HOLLAND, V.L. 1988. Coastal oak woodland. Pages 78-79
in K.E. Mayer and W.F. Laudenslayer, Jr. [EDs.], A
guide to wildlife habitats of California. USDA Forest
Service, Sacramento, CA U.S.A.
INTERGRAPH CORPORATION. 1994. MGE base mapper ref-
erence manual. Intergraph, Huntsville, AL U.S.A.
JOLLY, G.M. 1965. Explicit estimates from capture-recap-
ture data with both death and immigration-stochastic
model. Biometrika 52:225-247.
1982. Mark-recapture models with parameters
constant in time. Biometrics 38:301-321.
KI•BS, C.J. 1985. Ecology: the experimental analysis of
distribution and abundance. Harper and Row, New
York, NY U.S.A.
LAHAY•, W.S. AND RJ. GUTII•RREZ. 1994. Big Bear Spotted
Owl study, 1993. Calif. Dept. of Fish and Game, Non-
game Bird and Mammal Section. Tech. Rep. 1994-3,
Sacramento, CA U.S.A.
--AND --'. 1999. Nest sites and nesting habitat
of the Northern Spotted Owl in northwestern Cah-
fornia. Condor 101:324-330.
LEBKETON, J.-D., K.P. BURNH^M, J. CLOBERT AND D.R. AN-
DERSON. 1992. Modeling survival and testing biologi-
cal hypotheses using marked animals: a unified ap-
proach with case studies. Ecol. Monogr. 62:67-118.
MARCOT, B.G. ANDJ. GARDETTO. 1980. Status of the Spot-
ted Owl in Six Rivers National Forestß West. Birds 11'
79-87ß
--AND J.W. THOMAS. 1997. Of Spotted Owls, old
growth, and new policies: a history since the Inter-
agency Scientific Committee Reportß USDA For. Serv.
Gen. Tech. Rep. PNW-408, Portland, OR UßSßAß
MA•ER, K.E. 1988. Redwoodß Pages 60-61 in K.E. Mayer
and W.F. Laudenslayer, Jr. [EDs.], A guide to wildlife
habitats of California, USDA Forest Service, Sacra-
mento, CA U.S.A.
MCCULLAGH, P. AND J.A. NELDER. 1989. Generalized hn-
ear models. Chapman and Hall, New York, NY U.S.A.
MOEN, C.A., A.B. FP,2',NKLIN AND RJ. GUTIgRP, EZ. 1991. Age
determination of subadult Northern Spotted Owls tn
northwest California. Wildl. Soc. Bull. 19:489-493.
--AND R.J. GUTII•RREZ. 1993. Population ecology of
the California Spotted Owl in the central Sierra Ne-
vada: annual results, 1992. Calif. Dept. of Fish and
Game, Nongame Bird and Mammal Section. Tech.
Rep. 1993-14, Sacramento, CA U.S.A.
NETER, J. AND W. WASSERMAN. 1974. Applied linear staus-
tical modelsß Richard D. Irwin, Homewood, IL U.S.A
ODUM, E.P. 1971. Fundamentals of ecology. Saunders Col-
lege Publ., Philadelphia, PA U.S.Aß
POLLOCK, K.H.,J.E. HINES ANDJ.D. NICHOLS. 1985. Good-
ness-of-fit tests for open capture-recapture models
Biometrics 41: 399-410.
, J.D. NICHOLS, C. BROWNIE AND J.E. HINES. 1990.
Statistical inference for capture-recapture experi-
ments. Wildl. Monogz. 107:1-97.
ROBERTS, C.K. 1993. California Spotted Owl (Strix occiden-
talis caurina) inventory and demographic study, Se-
quoia and Kings Canyon National Parks: final 1988-
89. Calif. Dept. of Fish and Game, Nongame Bird and
Mammal Section. Tech. Rep. 1993-4, Sacramento, CA
U.S.A.
SAUER, RJ. AND B.K. WILLIAMS. 1989. Generalized pro-
cedures for testing hypotheses about survival or re-
covery rates. J. Wildl. Manage. 53:137-142.
SEBER, G.A.E 1965. A note on the multiple recapture cen-
sus. Biometrika 52:249-259.
ß 1982. The estimation of animal abundance and
related parameters. Charles Griffin and Co., London,
U.K.
SIMPSON TIMBER COMPANY. 1992. Habitat conservauon
plan for the Northern Spotted Owl on the Califorma
timberlands of Simpson Timber Company. Simpson
Timber Company, Korbel, CA U.S.A.
SOLIS, D.M., JR. AND RJ. GUTII•RREZ. 1990. Summer hab-

286 D•LI•E}• AND THOME VOL. 33, NO. 4
itat ecology of Northern Spotted Owls in northwest-
ern California. Condor 92:739-748.
TaN•4E}•, R.G. HI) RJ. GUTI•P,J•Z. 1995. A partial inven-
tory of Northern Spotted Owls (Strix occidentalis caur-
ina) in Redwood National Park, 1994. Unpubl. Rep.,
Arcata, CA U.S.A.
THOM^S, J.W., E.D. FORSMAN, J.B. LINT, E.C. MESI•OW,
B.R. NOO•4 aNI)J. VEP, NE}•. 1990. A con'servation strat-
egy for the Northern Spotted Owl. U.S. Gov. Printing
Off., Washington, DC U.S.A.
THOME, D.M., C.J. Z•mEL aND L.V. DILLEP,. 1999. Forest
stand characteristics and reproduction of Spotted
Owls in managed north-coastal California forests. J.
Wildl. Manage. 63:44-59.
USDA. 1995. Land and resource management plan for
the Six Rivers National Forest: final environmental im-
pact statement. Eureka, CA U.S.A.
USDI. 1992. Draft recovery plan for the Northern Spot-
ted Owl. USDI Fish and Wildl. Serv., Portland, OR
U.S.A.
V^N HORNE, B. 1983. Density as a misleading indicator
of habitat quality. J. Wildl. Manage. 47:893-901.
ZINI•, PJ. 1988. The redwood forest and associated north
coast forests. Pages 679-698 in M.G. Barbour and J.
Major [EI)s.], Terrestrial vegetation of California.
Univ. of Calif. Davis, Davis, CA U.S.A.
Received 28 January 1999; accepted 24 July 1999
